Glioblastoma (GBM) is a kind of intractable brain tumor. The effect of surgical treatment, radiotherapy and chemotherapy is not ideal. TRF1 is one of the important components of shelterin complex, which plays an important role in human telomere protection. Previous studies have reported that inhibition of TRF1 expression can inhibit the growth and proliferation of GBM without causing serious physiological dysfunction. However, the specific mechanism of inhibition of GBM growth and proliferation caused by decreased TRF1 expression has not been fully elucidated. To further elucidate this mechanism, we knockdown TRF1 by siRNA. We detected the levels of cell senescence, autophagy through biological experiments. It has been found that the knockdown of TRF1 can cause significant increase in the aging, autophagy of GBM. In addition, SIRT-6 is a NAD+- dependent deacetylase. Previous studies have reported that SIRT-6 can maintain the stability of telomere function. Moreover, telomere dysfunction can cause the decrease of SIRT-6 expression. Therefore, we want to study the effect of SIRT-6 expression level on TRF1 knockdown induced aging, autophagy in GBM. The experimental results showed that the knockdown of TRF1 caused the decrease of SIRT-6 expression level, and the increase of SIRT-6 expression level could inhibit the aging, autophagy caused by TRF1 knockdown. This study provides a new direction for the treatment of GBM.
| Published in | International Journal of Biomedical Engineering and Clinical Science (Volume 11, Issue 2) |
| DOI | 10.11648/j.ijbecs.20251102.11 |
| Page(s) | 11-27 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Glioblastoma, TRF1, SIRT-6, Autophagy, Senescence, Telomere Damage
Sence | Antisence | |
|---|---|---|
TRF1-siRNA1 | GGUGAUCCAAAUUCUCAUATT | UAUGAGAAUUUGGAUCACCTT |
TRF1-siRNA2 | GGUACAGUAUCCUUAUUGATT | UCAAUAAGGAUACUGUACCTT |
Negative control (NC) | UUCUCCGAACGUGUCACGUTT | ACGUGACACGUUCGGAGAATT |
Sence | Antisence | |
|---|---|---|
TRF1 | AACAGCGCAGAGGCTATTATTC | CCAAGGGTGTAATTCGTTCATCA |
p16 | GGAGGCCGATCCAGGTCAT | CACCAGCGTGTCCAGGAAG |
SIRT-6 | TCCCCGACTTCAGGGGTC | GTTCTGGCTGACCAGGAAGC |
p53 | AAGTCTAGAGCCACCGTCCA | CAGTCTGGCTGCCAATCCA |
ATG-5 | ATGATAATGGCAGATGACAAGG | TCAGTCACTCGGTGCAGG |
Beclin-1 | AGGTTGAGAAAGGCGAGACA | AATTGTGAGGACACCCAAGC |
IL-1α | GGTTGAGTTTAAGCCAATCCA | TGCTGACCTAGGCTTGATGA |
IL-6 | CAGGAGCCCAGCTATGAACT | GAAGGCAGCAGGCAACAC |
GAPDH | GTCTCCTCTGACTTCAACAGCG | ACCACCCTGTTGCTGTAGCCAA |
TRF1 | Telomere Repeat Sequence Binding Factor 1 |
TRF2 | Telomere Repeat Sequence Binding Factor 2 |
GBM | Glioblastomamultiforme |
SIRT6 | NAD-dependent Protein Deacetylase Sirtuin-6 |
SASP | Senescence-associated Secretory Phenotype |
ATM | Ataxia Telangiectasia Mutated |
IL-1α | Interleukin-1α |
IL-6 | Interleukin-6 |
LC3 | Microtubule-associatedprotein1 Light Chain 3 |
ATG-5 | Autophagy-relatedprotein-5 |
| [1] | Venkataramani, V. T. K. et al. Glioblastoma hijacks neuronal mechanisms for brain invasion. Cell |
| [2] | Dai L, Liu Z, Zhu Y, Ma L (2023b) Genome-wide methylation analysis of circulating tumor DNA: a new biomarker for recurrent glioblastom. Heliyon 9(3): e14339. |
| [3] | Fujita Y, Nunez-Rubiano L, Dono A, Bellman A, Shah M, Rodriguez JC, Putluri V, Kamal AHM, Putluri N, Riascos RF, Zhu JJ, Esquenazi Y, Ballester LY (2022) IDH1 p. R132H ctDNA and D-2-hydroxyglutarate as CSF biomarkers in patients with IDH-mutant gliomas. J Neuro-oncol 159(2): 261–270. |
| [4] | Brankiewicz W, Kalathiya U, Padariya M et al (2023) Modified peptide molecules as potential modulators of shelterin protein functions; TRF1. Chem – Eur J 29: e202300970. |
| [5] | Kameron Azarm, Amit Bhardwaj, Eugenie Kim and Susan Smith,“Persistent telomere cohesion protects aged cells from premature senescence, ”NATURE COMMUNICATIONS (2020) 11: 3321 | |
| [6] | Chen X, Tang W-J, Shi JB et al (2020) Therapeutic strategies for targeting telomerase in cancer. Med Res Rev 40: 532–585. |
| [7] | Ruis P, Boulton SJ. The end protection problem—an unexpected twist in the tail. Genes Dev. 2021. Jan 1; 35(1–2): 1–21. |
| [8] | Smith EW, Lattmann S, Liu ZB, Ahsan B, Rhodes D. Insights into POT1 structural dynamics revealed by cryo-EM. PLOS ONE. 2022. Feb 17; 17(2): e0264073. |
| [9] | Heyza JR, Mikhova M, Bahl A, Broadbent D, Schmidt JC. eLife. eLife Sciences Publications Limited; 2023. [cited 2023 Jul 11]. Systematic analysis of the molecular and biophysical properties of key DNA damage response factors. |
| [10] | Kim GJ, Lee JH, Chae M, et al. Prognostic value of telomeric zinc finger-associated protein expression in adenocarcinoma and squamous cell Carcinoma of lung. Medicina. 2021; 57(11): 1223. |
| [11] | Weiss F., Lauffenburger D., Friedl P. Towards Targeting of Shared Mechanisms of Cancer Metastasis and Therapy Resistance. Nat. Rev. Cancer. 2022; 22: 157–173. |
| [12] | Yamamoto H., Matsui T. Molecular Mechanisms of Macroautophagy, Microautophagy, and Chaperone-Mediated Autophagy. J. Nippon Med. Sch. 2023; 91: 2–9. |
| [13] | Miwa S, Kashyap S, Chini E et al. Mitochondrial dysfunction in cell senescence and aging. J Clin Invest. 2022; 132: e158447. |
| [14] | Oppedisano F, Nesci S, Spagnoletta A. Mitochondrial sirtuin 3 and role of natural compounds: the effect of post-translational modifications on cellular metabolism. Critical Reviews in Biochemistry and Molecular Biology. 2024; 59: 199–220. |
| [15] | Wu Q-J, Zhang T-N, Chen H-H, Yu X-F, Lv J-L, Liu Y-Y. et al. The sirtuin family in health and disease. Sig Transduct Target Ther. 2022; 7: 402. |
| [16] | Wu YX, Xu RY, Jiang L, Chen XY, Xiao XJ. Microrna‐30a‐5p promotes chronic heart failure in rats by targeting sirtuin‐1 to activate the nuclear factor‐κb/nod‐like receptor 3 signaling pathway. Cardiovasc Drugs Ther. 2022; 37: 1065‐1076. |
| [17] | Liu G., Chen H., Liu H., Zhang W., and Zhou J., “Emerging Roles of SIRT6 in Human Diseases and Its Modulators,” Medicinal Research Reviews 41 (2021): 1089–1137. |
| [18] | Protsenko E., Rehkopf D., Prather A. A., Epel E., Lin J. Are long telomeres better than short? Relative contributions of genetically predicted telomere length to neoplastic and non-neoplastic disease risk and population health burden. PLoS ONE. 2020; 15: e0240185. |
| [19] | Osum M., Serakinci N. Impact of circadian disruption on health; SIRT1 and Telomeres. DNA Repair. 2020; 96: 102993. |
| [20] | Haag, D. et al. H3.3-K27M drives neural stem cell-specific gliomagenesis in a human iPSC-derived model. Cancer Cell39, 407–422. e13 (2021). |
| [21] | Schaff LR, Mellinghoff IK. Glioblastoma and other primary brain malignancies in adults: A review. JAMA. 2023; 329(7): 574–87. |
| [22] | Rodrigues J, Alfieri R, Bione S et al. TERRA ONTseq: a long-read-based sequencing pipeline to study the human telomeric transcriptome. RNA. 2024; 30: 955–66. |
| [23] | Deng L., Yao P., Li L., Ji F., Zhao S., Xu C., Lan X., Jiang P. p53-mediated control of aspartate-asparagine homeostasis dictates LKB1 activity and modulates cell survival. Nat Commun. 2020; 11(1): 1755. |
| [24] | Yang Y, Ning C, Li Y, Wang Y, Hu J, Liu Y, et al. Dynamic changes in mitochondrial DNA, morphology, and fission during oogenesis of a seasonal-breeding teleost, Pampus argenteus. Tissue Cell. 2021; 72: 101558. |
| [25] | Smeriglio P, Zalc A. Cranial neural crest cells contribution to craniofacial bone development and regeneration. Curr Osteoporos Rep. 2023; 21: 624–3. |
| [26] | Singh V, Ubaid S. Role of Silent Information Regulator 1 (SIRT1) in Regulating Oxidative Stress and Inflammation. Inflammation (2020) 43(5): 1589–98. |
| [27] | Porreca RM, Herrera-Moyano E, Skourti E, Law PP, Gonzalez Franco R, Montoya A, Faull P, Kramer H, Vannier J-B. 2020. TRF1 averts chromatin remodelling, recombination and replication dependent-break induced replication at mouse telomeres. Elife 9: 1–28. |
| [28] | Hayes JD, Dinkova-Kostova AT, Tew KD Oxidative stress in cancer. Cancer Cell. 2020; 38: 167–97. |
| [29] | Kim AJ, Xu N, Yutzey KE. Macrophage lineages in heart valve development and disease. Cardiovasc Res. 2021; 117(3): 663–673. |
| [30] | Batshon G, et al. Serum NT/CT SIRT1 ratio reflects early osteoarthritis and chondrosenescence. Ann. Rheum. Dis. 2020; 79: 1370–1380. |
| [31] | Wang L., Klionsky D. J., Shen H. M. The Emerging Mechanisms and Functions of Microautophagy. Nat. Rev. Mol. Cell Biol. 2023; 24: 186–203. |
| [32] | Norberto Vargas J. S., Hamasaki M., Kawabata T., Youle R. J., Yoshimori T. The Mechanisms and Roles of Selective Autophagy. Nat. Rev. Mol. Cell Biol. 2023; 24: 167–185. |
| [33] | Deneubourg C, Ramm M, Smith LJ. The spectrum of neurodevelopmental, neuromuscular and neurodegenerative disorders due to defective autophagy. Autophagy. 2022; 18: 496‐517. |
| [34] | Karunarathne, W. et al. Anthocyanin-enriched polyphenols from Hibiscus syriacus L. (Malvaceae) exert anti-osteoporosis effects by inhibiting GSK-3beta and subsequently activating beta-catenin. Phytomedicine91, 153721 (2021). |
| [35] | Deng, Z. et al. Melatonin attenuates sepsis-induced acute kidney injury by promoting mitophagy through SIRT3-mediated TFAM deacetylation. Autophagy20, 151–165 (2024). |
| [36] | Jaewon Min, Woodring E. Wright, and Jerry W. Shay. Clustered telomeres in phase-separated nuclear condensates engage mitotic DNA synthesis through BLM and RAD52. GENES & DEVELOPMENT 33: 814–827; ISSN 0890-9369/19; |
| [37] | Paula Martı′nez, Maria Thanasoula, Purificacio′n Mun˜ oz, et al., “Increased telomere fragility and fusions resulting from TRF1 deficiency lead to degenerative pathologies and increased cancer in mice, ”GENES & DEVELOPMENT 23: 2060–2075; ISSN 0890-9369/09; |
| [38] | Lim CJ, Cech TR. Shaping human telomeres: from shelterin and CST complexes to telomeric chromatin organization. Nat Rev Mol Cell Biol. 2021; 22: 283–98. |
| [39] | Li Q, Ma Q, Xu L, Gao C, Yao L, Wen J, Yang M, Cheng J, Zhou X, Zou J, Zhong X, Guo X. Human Telomerase Reverse Transcriptase as a Therapeutic Target of Dihydroartemisinin for Esophageal Squamous Cancer. Front Pharmacol. 2021; 12: 769787. |
| [40] | Jayaram M. A., Phillips J. J. Role of the Microenvironment in Glioma Pathogenesis. Annu. Rev. Pathol. 2024; 19: 181–201. |
| [41] | Liu Q, Liu Y, Li SE, Geng JH. Bcl-2 interacts with beclin 1 and regulates autophagy in 7, 12-dimethylbenz[a]anthracene-induced hamster buccal-pouch squamous-cell tumorigenesis. Curr Med Sci. (2021) 41: 1198–204. |
APA Style
Nie, Z., Liu, S., Huang, X., Geng, X. (2025). Inhibition of TRF1 Can Accelerate Aging and Induce Autophagy Through the P53-SIRT6 Pathway in Glioblastoma Multiforme. International Journal of Biomedical Engineering and Clinical Science, 11(2), 11-27. https://doi.org/10.11648/j.ijbecs.20251102.11
ACS Style
Nie, Z.; Liu, S.; Huang, X.; Geng, X. Inhibition of TRF1 Can Accelerate Aging and Induce Autophagy Through the P53-SIRT6 Pathway in Glioblastoma Multiforme. Int. J. Biomed. Eng. Clin. Sci. 2025, 11(2), 11-27. doi: 10.11648/j.ijbecs.20251102.11
AMA Style
Nie Z, Liu S, Huang X, Geng X. Inhibition of TRF1 Can Accelerate Aging and Induce Autophagy Through the P53-SIRT6 Pathway in Glioblastoma Multiforme. Int J Biomed Eng Clin Sci. 2025;11(2):11-27. doi: 10.11648/j.ijbecs.20251102.11
@article{10.11648/j.ijbecs.20251102.11,
author = {Ziyang Nie and Siqi Liu and Xinyu Huang and Xin Geng},
title = {Inhibition of TRF1 Can Accelerate Aging and Induce Autophagy Through the P53-SIRT6 Pathway in Glioblastoma Multiforme
},
journal = {International Journal of Biomedical Engineering and Clinical Science},
volume = {11},
number = {2},
pages = {11-27},
doi = {10.11648/j.ijbecs.20251102.11},
url = {https://doi.org/10.11648/j.ijbecs.20251102.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijbecs.20251102.11},
abstract = {Glioblastoma (GBM) is a kind of intractable brain tumor. The effect of surgical treatment, radiotherapy and chemotherapy is not ideal. TRF1 is one of the important components of shelterin complex, which plays an important role in human telomere protection. Previous studies have reported that inhibition of TRF1 expression can inhibit the growth and proliferation of GBM without causing serious physiological dysfunction. However, the specific mechanism of inhibition of GBM growth and proliferation caused by decreased TRF1 expression has not been fully elucidated. To further elucidate this mechanism, we knockdown TRF1 by siRNA. We detected the levels of cell senescence, autophagy through biological experiments. It has been found that the knockdown of TRF1 can cause significant increase in the aging, autophagy of GBM. In addition, SIRT-6 is a NAD+- dependent deacetylase. Previous studies have reported that SIRT-6 can maintain the stability of telomere function. Moreover, telomere dysfunction can cause the decrease of SIRT-6 expression. Therefore, we want to study the effect of SIRT-6 expression level on TRF1 knockdown induced aging, autophagy in GBM. The experimental results showed that the knockdown of TRF1 caused the decrease of SIRT-6 expression level, and the increase of SIRT-6 expression level could inhibit the aging, autophagy caused by TRF1 knockdown. This study provides a new direction for the treatment of GBM.
},
year = {2025}
}
TY - JOUR T1 - Inhibition of TRF1 Can Accelerate Aging and Induce Autophagy Through the P53-SIRT6 Pathway in Glioblastoma Multiforme AU - Ziyang Nie AU - Siqi Liu AU - Xinyu Huang AU - Xin Geng Y1 - 2025/05/29 PY - 2025 N1 - https://doi.org/10.11648/j.ijbecs.20251102.11 DO - 10.11648/j.ijbecs.20251102.11 T2 - International Journal of Biomedical Engineering and Clinical Science JF - International Journal of Biomedical Engineering and Clinical Science JO - International Journal of Biomedical Engineering and Clinical Science SP - 11 EP - 27 PB - Science Publishing Group SN - 2472-1301 UR - https://doi.org/10.11648/j.ijbecs.20251102.11 AB - Glioblastoma (GBM) is a kind of intractable brain tumor. The effect of surgical treatment, radiotherapy and chemotherapy is not ideal. TRF1 is one of the important components of shelterin complex, which plays an important role in human telomere protection. Previous studies have reported that inhibition of TRF1 expression can inhibit the growth and proliferation of GBM without causing serious physiological dysfunction. However, the specific mechanism of inhibition of GBM growth and proliferation caused by decreased TRF1 expression has not been fully elucidated. To further elucidate this mechanism, we knockdown TRF1 by siRNA. We detected the levels of cell senescence, autophagy through biological experiments. It has been found that the knockdown of TRF1 can cause significant increase in the aging, autophagy of GBM. In addition, SIRT-6 is a NAD+- dependent deacetylase. Previous studies have reported that SIRT-6 can maintain the stability of telomere function. Moreover, telomere dysfunction can cause the decrease of SIRT-6 expression. Therefore, we want to study the effect of SIRT-6 expression level on TRF1 knockdown induced aging, autophagy in GBM. The experimental results showed that the knockdown of TRF1 caused the decrease of SIRT-6 expression level, and the increase of SIRT-6 expression level could inhibit the aging, autophagy caused by TRF1 knockdown. This study provides a new direction for the treatment of GBM. VL - 11 IS - 2 ER -
Department of Biochemistry and Molecular Biology, School of Basic Medical Sciences, Tianjin Medical University, Tianjin, ChinaKey Laboratory of Immune Microenvironment and Disease (Ministry of Education), Tianjin Medical University, Tianjin, China Key Laboratory of Immune Microenvironment and Disease (Ministry of Education), Tianjin Medical University, Tianjin, China School of Life Sciences, Central China Normal University, Wuhan, China
Department of Biochemistry and Molecular Biology, School of Basic Medical Sciences, Tianjin Medical University, Tianjin, China Key Laboratory of Immune Microenvironment and Disease (Ministry of Education), Tianjin Medical University, Tianjin, China
Department of Biochemistry and Molecular Biology, School of Basic Medical Sciences, Tianjin Medical University, Tianjin, China Key Laboratory of Immune Microenvironment and Disease (Ministry of Education), Tianjin Medical University, Tianjin, China
Department of Biochemistry and Molecular Biology, School of Basic Medical Sciences, Tianjin Medical University, Tianjin, China Key Laboratory of Immune Microenvironment and Disease (Ministry of Education), Tianjin Medical University, Tianjin, China
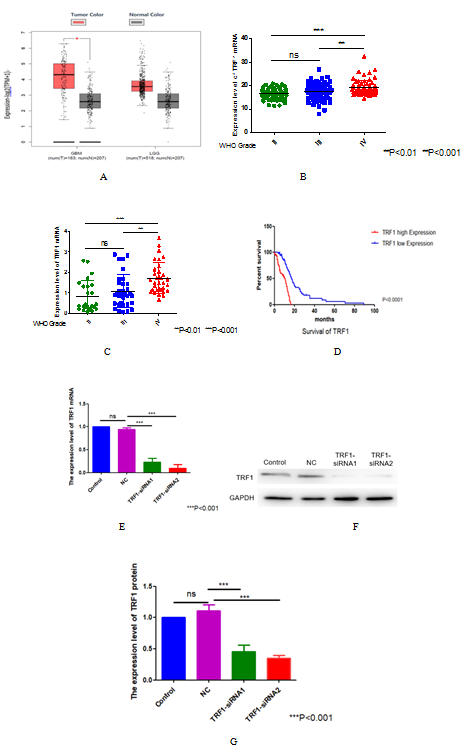
Figure 1. The expression level of TRF1 in glioma and the knockdown efficiency of TRF1. (A) The expression of TRF1 mRNA in glioma was analyzed by GEPIA database. (B) The expression levels of TRF1 mRNA in different grades of gliomas were analyzed by CCGA database. (C) The expression levels of TRF1 mRNA in different grades of gliomas were analyzed by TCGA database. (D) ATLAS database speculates the relationship between TRF1 mRNA expression and prognosis of glioma patients. The subjects of the statistics are all IDH wild-type glioma patients. (E) The knockdown efficiency of TRF1 in U87 cells was detected by qRT-PCR. GAPDH was used as internal control. (F) The knockdown efficiency of TRF1 was detected by Western blot. (G) Quantitative analysis of (D). ***P<0.001.
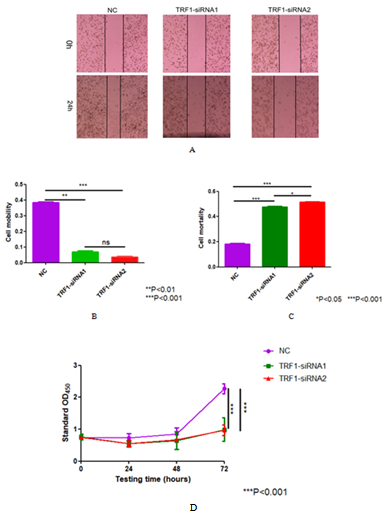
Figure 2. Effect of knocking down TRF1 on the growth and proliferation of U87 cells. (A) Scratch test was used to detect the effect of TRF1 knockdown on U87 cell migration. (B) Quantitative analysis of (A). (C) Trypan blue assay was used to detect the effect of TRF1 knockdown on the survival status of U87 cells. (D) CCK-8 assay was used to detect the effect of TRF1 knockdown on the proliferation of U87 cells.*P<0.05 **P<0.01 ***P<0.001.
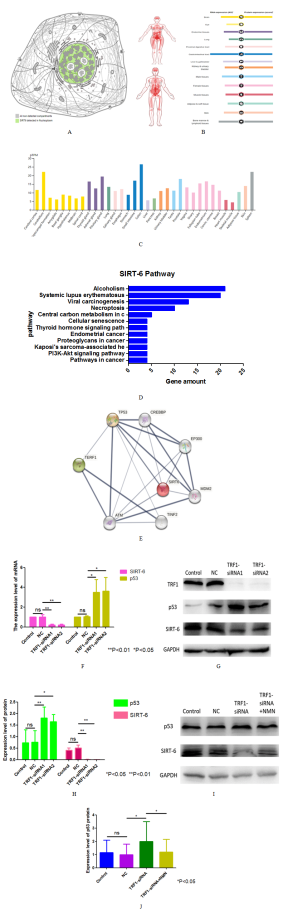
Figure 3. The relationship between TRF1, p53 and SIRT-6. (A) Distribution of SIRT-6 in glioma cells analyzed by Atlas Database. (B-C) Protein and mRNA expression of SIRT-6 in human tissues. (D) The relationship among TRF1, p53 and sirt-6 was inferred by String database. (E-F) The mRNA and protein expression levels of sirt-6 and p53 in TRF1 knockdown cells were detected. (G) The quantitative analysis of (H) The expression of SIRT-6 and p53 protein in U87 cells added with NMN after TRF1 knockdown was detected by Western blot. The final concentration of NMN was 250uM. (I) The quantitative analysis of (I). *P<0.05 **P<0.01.
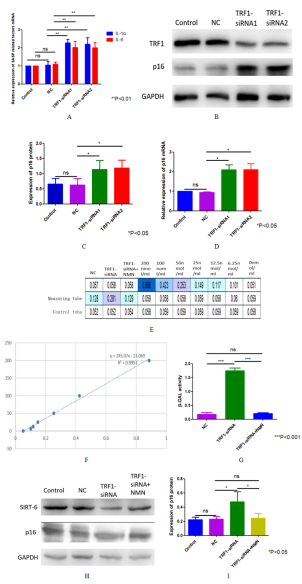
Figure 4. Effect of knocking down TRF1 on senescence of U87 cells and its specific mechanism. (A) The mRNA expressions of IL-1α and IL-6 in TRF1 knockdown cells were detected by qRT-PCR. (B-D) Western blot and qRT-PCR were used to detect the expression of p16 protein and mRNA in the TRF1 knockdown cells. (E) β- Gal activity (UV spectrophotometry) was used to detect the aging status of TRF1 knockdown and NMN added after TRF1 knockdown. (F) Standard curve of β-GAL activity experiment. (G) Quantitative analysis of β-GAL activity. (H) Western blot was used to detect the p16 expression of TRF1 knockdown and NMN after TRF1 knockdown. (I) Quantitative analysis of (H). *P<0.05 **P<0.01 ***P<0.001.
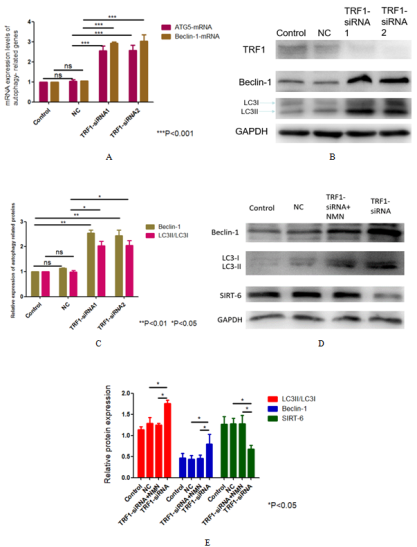
Figure 5. Mechanism of autophagy induced by knocking down TRF1 in U87 cells. (A, B) The mRNA and protein expression levels of autophagy markers after TRF1 knockdown were detected by qRT-PCR and Western blot. (C) Quantitative analysis of (B) (D) TRF1 knockdown and the changes of autophagy markers expression caused by adding NMN were detected by Western blot. (E) Quantitative analysis of (D). *P<0.05 **P<0.01 ***P<0.001.
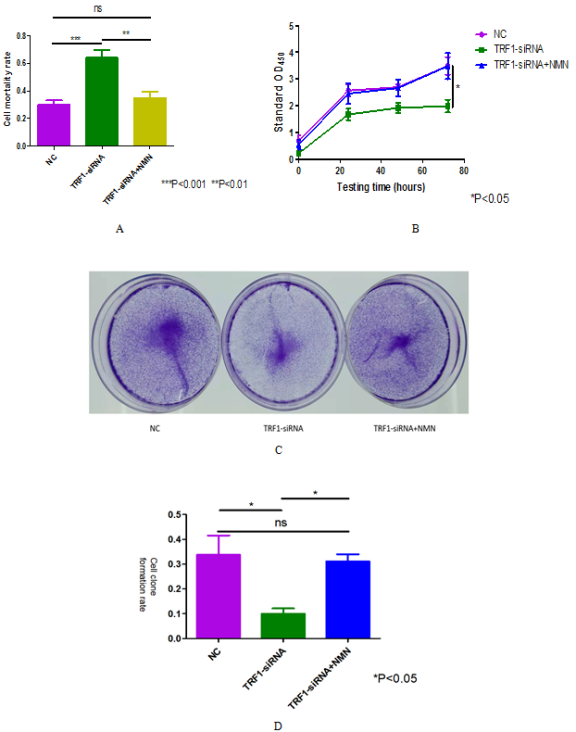
Figure 6. SIRT-6 can reduce the growth and proliferation restriction of U87 cells caused by TRF1 knockdown. (A) The death number of U87 cells in each experimental group was detected by trypan blue staining. (B) The ultraviolet absorbance at 450nm of U87 cells in each experimental group was detected by CCK-8 experiment. (C) The growth and proliferation of U87 cells in each experimental group were detected by cell clone formation experiment. (D) Quantitative analysis of (C).*P<0.05 **P<0.01 ***P<0.001.

